Xerocomellus sarnarii Simonini, Vizzini & Eberhardt
Index Fungorum number: IF551434 Facesoffungi number: FoF01047
Etymology: dedicated to our friend Mauro Sarnari, who found this bolete at Monte Argentario.
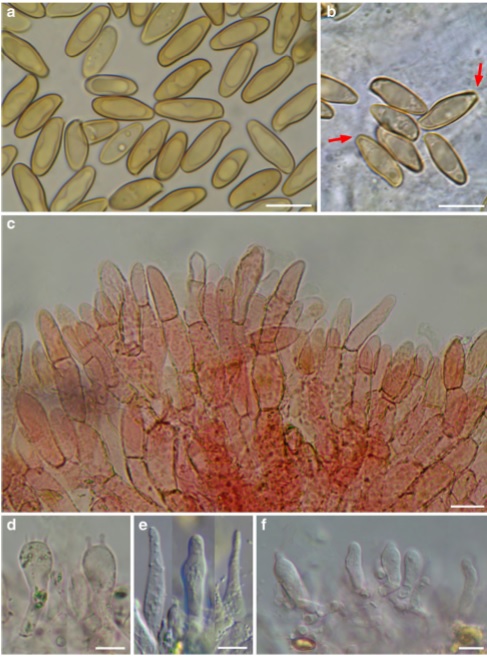
Holotype: MCVE 28577 Colour terminology and alphanumeric codes are those of Kornerup and Wanscher (1978) and Seguy (1936). Pileus 35–90 mm broad, fleshy, convex to pulvinate, rarely almost plane, also slightly depressed at the centre but conserving pulvinate shape; surface dry, tomentose-velutinous, soon cracking into scabs independently on weather conditions, with a subpellis very weakly and not always pink-reddish coloured, being this colour rarely visible in the fissures, sometimes only in the bites of the snails; surface variable in colour: in very young basidiomes with uncracked pileus dark brown with olive shade (6–7/F6–F4, 6/F8–F7), with fading patches (5/D5–D3), cracking into scabs and discolouring to ochrebeige with olive shade (4–5/D6–D3), passing through transition colours (5–6/D7–F4). Dull cream-whitish (3/4A2) cracks appear beneath the pileipellis layer in dry weather. Tubes up to 5–10 mm long, separable, depressed to sinuate around the stipe, sometimes slightly decurrent, as high as the context of the pileus or even more; initially bright yellow with olive shades (1/2A7/8, 3A/B7/8), later dull greenish yellow (2C/D7/8, 3/4C/D6), bluing then darkening to blackish when bruised. Pores angular, uneven, in mature basidiomes 0.7– 2 mm diam., concolourous to the tubes, slowly bluing (S351, 352, 366, 367) then darkening to blackish when bruised. Stipe 30–90×5–25 mm, cylindrical, slender, curved, somewhat tapering towards the base or also slightly widened. Surface dry, smooth to ingrown fibrillose, chrome yellow in the upper part, in the lower part and towards the base or partially or completely, sometimes abruptly, red, dark purple red or even blackish-red coloured (11–12/D8–F4). Context soft in the pileus, fibrillose and brittle in the stipe; in the pileus pale yellow (3A3) soon fading to dirty whitish when exposed (29/ 30A2), below the pileipellis sometimes showing a fine red line, often inconspicuous or really absent; in the stipe vivid yellow (3A4/5) or yellow with dull reddish tinges that are concolourous to the cortex (8 to 11C/D7/8), bluing (S446 to 448) in the whole pileus and in the stipe upper part when cut, with the dark purple red areas in the stipe base darkening, the stipe context at the extreme base dull ochre-brownish (S336 to 339); smell not distinctive, typical of the Xerocomellus, somewhat like wet iron; taste not distinctive, slightly sour. Sporeprint: brownish with some olive tinges. Macrochemical reactions: weak fleeting-amyloid reaction in stipe context and hymenophoral trama of dry specimens. Spores (13.5) 13.8–15.1 (15.7)× (5.5) 5.5–6.1 (6.6) μm, Q=2.51±0.08, V=256±41 μm3 (n=330/10/10), subfusiform, slender, with a weakly developed but distinct supra-apicular depression, spore wall up to 0.5 μm thick, intensely honey-coloured and with one or two guttules when mature, with a specialized spore apex (“truncature” faint and not always visible under the optical MS). Basidia 25–36×10–14 μm, inconspicuous, clavate, hyaline to yellowish, mainly 4-spored, not rarely 2- spored. Pleurocystidia scarce, 35–52×6–11 μm, fusiform or ventricose-fusiform, sometimes with a obtuse-rounded apex, resembling those of Psathyrella barlae (Bres.) A.H. Sm., with hyaline to slightly yellowish content. Cheilocystidia similar, but smaller, 22–32×4–10. In some of the specimens collected it was observed an abundance of pseudocystidia with a yellowish, refringent, oily content, reacting to dark blue with Cresyl blue. Pileipellis a palisadoderm strongly reminiscent of the pileipellis of X. chrysenteron, with long parallel chains of elements, terminal elements rarely wider than the basal ones, mostly cylindrical and somewhat tapering towards the apex, obtusely triangular, mitriform, also acorn-shaped to exceptionally subsphaerical, rarely papillate, (36.5) 36.2–38.2 (39.2)×(13.1) 13.3–15.9 (16.1) μm, Q=2.62±0.22 (n=99/3/3), terminal and subterminal elements moderately to heavily incrusted, only exceptionally smooth. Stipitipellis consisting of loosely interwoven, 3–6 μm broad hyphae with hyaline to yellowish content. In the upper part of the stipe is present a well-developed caulohymenium with spore-forming caulobasidia and caulocystidia. Stipe texture formed of parallel running, hyaline, 2.5–5 μm broad hyphae; no “pruinatushyphae” in the lower part of the stipe are present (Ladurner and Pöder 2000). No clamp-connections observed.
Microchemical reactions: with the exception of the amyloid reacting hyphae in the stipe, no particular reproducible macrochemical reactions were observed.
Habitat and known distribution: X. sarnarii is a rare species, so far known only from Italy (Sardinia and Tuscany) and France (see notes), in Mediterranean areas under Quercus suber and Quercus ilex. This taxon seems to prefer climatically moderate zones and undisturbed habitats. The main fruiting period is late autumn (November).
Material examined: ITALY, Sardinia, Province of OlbiaTempio Pausania, Common of Calangianus, along road SP 38 “Via S. Antonio”, loc. Monti di La Jescia, “Cava Merzari”, 40°56′21″N, 9°12′35″E, 425 m a.s.l., on granitic soil, under Quercus suber, mixed with Xerocomus cisalpinus, 10 November 2011, leg. G. Simonini (MCVE 28577, holotype), (GS10025, isotype); same locality and date of the typus, MCVE 28571, MCVE 28572, MCVE 28573, MCVE 28576, MCVE 28578, MCVE 28579; Sardinia, NU, Dorgali, Cala Gonone, “Via dei Lecci”, 40°16′25″ N, 9°36′37″ E, 230 m a.s.l., on calcareous soil, under Quercus ilex, 7 November 1994, leg. G. Redeuilh (MCVE 17797); Tuscany, GR, Monte Argentario, “Via Acquedotto Leopoldino”, 42°24′53″N, 11°10′1″E, 185 m a.s.l., on calcareous soil, under Quercus ilex, 1 November 1998, leg. M. Sarnari (MCVE 18361). Additional material examined. Xerocomellus cisalpinus: ITALY, Sardinia, Province of Olbia-Tempio Pausania, Common of Calangianus, along road SP 38 “Via S. Antonio”, loc. Monti di La Jescia, “Cava Merzari”, 40°56′21″N, 9°12′35″E, 425 m a.s.l., on granitic soil, under Quercus suber, mixed with Xerocomellus sarnarii, 10 November 2011, leg. G. Simonini (GS10020). Xerocomellus porosporus: ITALY, Latium, Manziana (RM), 320 m a.s.l., under Quercus cerris in acid soil, 20 June 2009, legit. M. Gelardi and V. Migliozzi (MG197a).
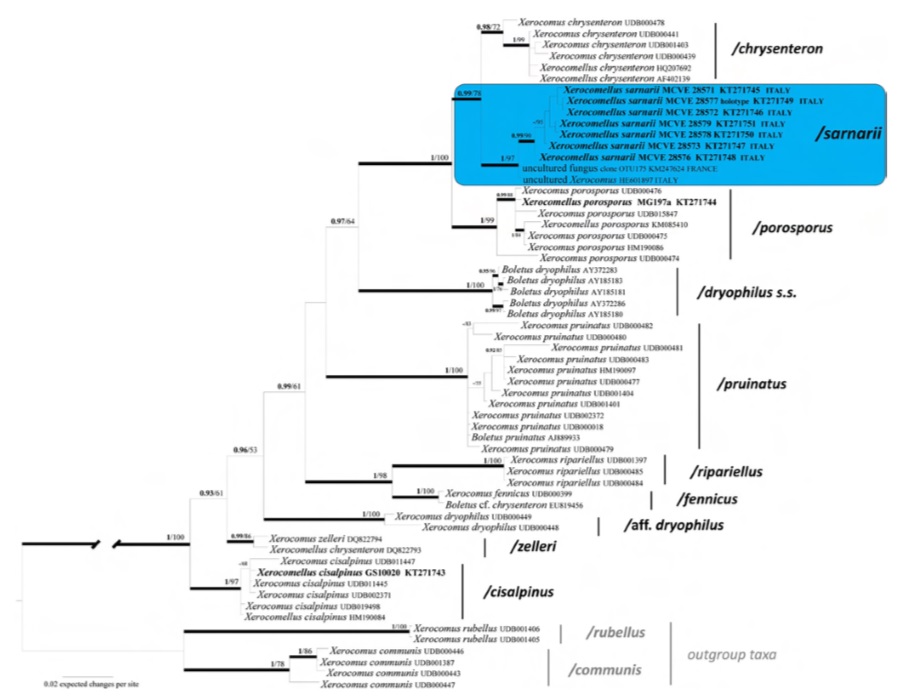
Notes: Both Bayesian and Maximum likelihood analyses produced the same topology; therefore, only the Bayesian tree with both BPP and MLB values is shown. In the ITS sequence analysis the 7 sequenced collections of X. sarnarii form a well-supported clade / sarnarii, (BPP=1,MLB=97 %), together with two environmental sequences (uncultured fungus clone OTU175 KM247624 from ectomycorhiza of Quercus ilex, Pézilla-de-Conflent, southern France, and uncultured Xerocomus HE601897 from ectomycorrhizal mantle of Quercus ilex, Sardinia, Italy). As the sequences of this clade show a high ITS sequence homology (pairwise % identity value=99.4), X. sarnarii seems present also in France. The X. sarnarii clade is sister (BPP=99, MLB=78 %) to X. chrysenteron. As with the other Xerocomellus species, X. sarnarii is difficult to be recognized in the field based only on its macromorphological features. At a first glance, the brownish cracked pileus with the pallid context showing through, recalls X. porosporus, but the vivid purple-red stipe base is disconcerting. In young specimens, or specimens in which the pileus surface does not crack clearly, the confusion with X. redeuilhii nom. prov. (Taylor et al. 2012 = X. dryophilus sensu Simonini 1994, and as X. aff. dryophilus) is possible, due to the two-coloured stipe cortex, chrome yellow in the upper part and abruptly dark red in the lower part, very close to that of X. redeuilhii.; moreover, some collections of X. redeuilhii may also have occasionally a brown pileus. The spores of the two boletes are however, clearly different mainly in Q ratio: Q=2.1–2.4 for X. redeuilhii and Q=2.4–2.6 for X. sarnarii. In its most typical appearance, because of its obvious cracked pileus surface, X. sarnarii is somewhat similar to X. chrysenteron (Bull.) Šutara, X. cisalpinus (Simonini et al.) Klofac, and X. porosporus (Imler ex Bon & G. Moreno) Šutara (= Boletus marekii Šutara et Skála, unpublished data); moreover, all these boletes have a similar pileipellis structure, with long chains of parallel, incrusted elements (Ladurner and Simonini 2003). Nevertheless, X. sarnarii can easily be distinguished from these species based on its spore features: the smooth spores with the characteristic specialized apex, showing a hardly detectable, small “truncation” are typical for X. sarnarii, and are different from both those of X. chrysenteron (without any truncation), X. cisalpinus (with no truncation and faintly but clearly longitudinally striate) and X. porosporus (smooth, but with a much more evident truncation) (Ladurner and Simonini 2003; Šutara 2008). In addition, the spores of X. sarnarii are significantly broader (5.5– 6.1 μm) than those of both X. chrysenteron (4.7–5.4 μm) and X. cisalpinus (4.6–5 μm). However, X. porosporus has spores as broad (5.6–6.4 μm) as X. sarnarii (Supplementary Table 4). Xerocomellus truncatus (Singer et al.) Klofac (2011), based on Xerocomus truncatus Singer et al. (1960), is a North American taxon that shares with X. sarnarii the general appearance, with the tendency of the pileus surface to crack and the stipe having also red areas. It differs for “the (pileus) cracks pinkish-purple or rarely with the pallid flesh showing through”, while the context among pileus cracks of X. sarnarii is, oppositely, whitish pallid and very rarely pinkish; the context “in the central portion of the stipe red”, while in X. sarnarii the red colour is concentrated in the lower half or the bottom; the spores “with a truncateapplanate apex which is thicker-walled at the two apical angles and often with markedly thinned wall between these angles, there also frequently depressed to umbilicate depressed, a minority with the apex merely rounded”, while in X. sarnarii the spores apex is only faintly truncate, and in the most of the cases the truncation is not clearly perceptible at the optical MS. In the later illustration by Snell and Dick (1970, pl. 79), the truncature is really wide and sharply angled in the spore adaxial profile, with an estimable width on the drawing=3 μm; the spores profile of X. sarnarii, is quite different, with a truncation that is hardly perceptible, having an estimable width=1 μm.