Cantharellus eucalyptorum Buyck, Randrianjohany & V. Hofstetter
MycoBank: MB 813239 Facesoffungi number: FoF00990
Etymology: refers to its association with Eucalyptus robusta.
Diagnosis: Differs from the Malagasy C. tricolor principally by the much paler gill folds, and genetically by its closer relationship to C. tanzanicus.
Holotype: MADAGASCAR, Central Highlands, 25 km before reaching Antananarivo when returning from Andasibe, collected under E. robusta in hills along the RN2 before reaching the village of Ambanitsena, 6 February 2006, Buyck & V. Hofstetter 06.159 (PC0085037). Cap mostly 25-45(−60) mm diam., regular or with uneven, waving or undulating lobes, when young with strongly incurved margin becoming even or uplifted when old with a sunken center to strongly infundibiliformous; surface when young tomentose-granulose to pubescent, with age usually fragmenting in larger, appressed and concentrically arranged squamules, when wet smooth to velvety, colours varying from cream or ochre (3A2 to 4A3-5) to pale grayish brown (5C4) or even dull gray in the center. Hymenophore decurrent and not abruptly delimited from the sterile stipe surface, formed of well-developed gill folds, 2-4(6) mm high, starting out as very pale cream or off-white (at the most 3-4A2), but then developing pale yellowish tints (3-4A4) with age, forkings not abundant except closer to the cap margin, either completely smooth or frequently anastomosed in between gill folds. Stipe mostly distinctly shorter than the cap diam., more rarely quite slender and exceeding the cap diam., (3-)5-10 mm diam., and mostly (15)20-30(45) mm long, subcylindrical or slightly narrowing downwards, mostly off-white to creamish yellow and somewhat paler than the hymenophore, in the upper half often developing appressed squamae deposited in circular girdings, solid. Flesh firm and thick beneath the cap center, whitish, but showing distinct yellowing when cut or handled, particularly in the lower stipe half. Taste mild. Smell faint and not very typical. Spore print probably pale yellowish, insufficient for a good appreciation. Spores ellipsoid to narrowly ellipsoid, often slightly constricted in the central part and reniformous, measuring (7.5)7.8-8.33-8.8(9.4)× (4.2)4.5-4.91-5.3(5.4) μm, Q=(1.5)1.6-1.71-1.8(1.9), neither amyloid nor cyanophilous, smooth, containing mostly a distinct, large oil drop. Basidia quite long and slender, mostly 50-70(−85)×5–7 μm, clavate, (2)4-5-spored. Cystidia absent. Subhymenial cells often long and slender, but often also wider than the basal part of the basidia. Lamellar trama comprised of predominantly narrow and slender elements, sometimes as thin as 1 μm diam., some being inflated towards the septa on one side. Pileipellis with thick-walled (up to 1 μm), flexuous hyphal extremities, ca 4– 9 μm wide, that often adhere together forming tufts or short, coiled, rhizoid-like structures, falling easily apart into smaller fragments when squashed, very frequently giving the impression that cells narrow abruptly near the septa and only stick together in the more central part of the septum; most cells slender and narrowly cylindrical; the terminal cell mostly 30–60 μm long, more often slightly inflated, clavate, fusoid or irregularly undulate in outline, near the tip obtuse-rounded or frequently constricted and papillate to subcapitate. Clamp connections absent from all tissues.
Material examined: MADAGASCAR, Central Highlands, Arivonimamo, under eucalypts, on lateritic soil at 1400 m alt., 30 January 1997, Buyck, Eyssartier & Moreau leg., in Buyck 97.078 (PC0084828), ibid., 30 January 1996, Buyck 96.543 (PC0084830); near Andasibe, under Eucalyptus, 12 February 1997, Buyck, Eyssartier & Moreau leg., in Buyck 97.406 (PC0084829); 25 km before reaching Antananarivo when returning from Andasibe, bought from vendors along the RN2 before reaching the village of Ambanitsena, 28 February 2000, Buyck 00.1828 (PC0085036); ibid. 4 February 2006, Buyck & V. Hofstetter V. 06.148 (PC0084127), 06.149 (PC0084128); ibid., collected under eucalypts, 6 February 2006, Buyck & V. Hofstetter 06.153 (PC0084112), 06.159 (PC0085037), 06.165 (PC0084789).
Notes: Cantharellus eucalyptorum is very closely related to the mainland African C. tanzanicus and we hesitated at first to describe it as a different species since our phylogeny shows no support to distinguish the two taxa, mainly due to the fact that C. tanzanicusis still known from a single collection. When taking a closer look at the sequences, the Malagasy specimens represent without any doubt a clearly different species. The 92 % BS that supports the distinction between a subclade formed by three specimens of C. eucalyptorum versus the grouping of the remaining two specimens together with C. tanzanicus, is based on a single mutation. However, the latter species differs from all six Malagasy specimens by four apomorphies in the coding part of the tef-1 sequences, and two additional mutations in the introns, which is far more than the number of apomorphies that support for example in subgenus Cantharellus the distinction between the North American C. tenuithrix, C. flavus and C. phasmatis (Foltz et al. 2013). Morphological characters that may help to distinguish C. eucalyptorum from C. tanzanicus are the slightly more voluminous spores, more pronounced filamentous subhymenium and somewhat longer basidia of the Malagasy species. Such differences, however, would need to be confirmed by comparing more collections. Judging from their general appearance, the specimens depicted and described in Härkönen et al. (2003, as C. isabellinus), most likely correspond to our C. tanzanicus and they share superposable spore sizes with the holotype of the latter [7.5–9×3.5–4.5 versus (7.3)7.9–8.4–8.9(9.2)× (3.7)3.8–4.1–4.4(4.6) μm], which suggests that the smaller and more cylindrical spores of C. tanzanicus might represent a reliable distinction. Spore measurements of other collections for C. eucalyptorum confirm those obtained for the holotype: 97.078/ (6.75)7-7.71-8.5(9)× (4.25)4.5-4.94-5.5(6) μm Q=(1.3)1.4-1.57-1.7(1.8) 96.543/ 7–7.37-7.7(8)×(4.5)4.8-5.17-5.5(6) Q=(1.3)1.5- 1.67-1.7 96.550/ (6.5)7.3-83-8.7(9.5)× (4)4.5-5.00-5.5(6) Q=(1.1)1.4-1.62-1.8(2.1) Holotype/ (7.5)7.8-8.33-8.8(9.4)×(4.2)4.5-4.91-5.3(5.4) μm, Q=(1.5)1.6-1.71-1.8(1.9) This fleshy and very variable chanterelle is locally abundant under E. robusta plantations on the Central Plateau of Madagascar and is frequently offered for sale along roads and in market places. It is often sold as‘girolle’, the common name in France for C. cibarius Fr.:Fr., of which it has the general habit, but not the yellow colour. It differs from this European species most markedly in the white, but distinctly yellowing flesh, particularly so in the stipe and, of course, in the grayish brown, fibrillose to cottony surface of the cap and to a lesser degree, also of the stipe. This type of surface is typical for many species in subgenus Rubrinus sect. Isabellinus (see Buyck et al. 2014), but can be quite variable in its appearance and development. This variability contributes a lot to the sometimes important differences in general aspect of these species. Typically smooth and continuous when young, at least in the cap center, this type of surface tissue frequently breaks up at maturity into concentric rings or into appressed to sometimes even dressed squamules that are more or less concentrically arranged. A similar pattern may be observed on the stipe surface with darker squamules in more or less horizontal arrangements circling the stipe, although the squamae are usually much less pronounced compared to the cap. The cap colour can vary from pale yellowish to quite dark brown or locally almost blackish gray when fresh and humid, but turns to an isabelline or dirty white or creamish colour when dry. The hymenophore is typically composed of gilllike folds that can be mixed with smaller lamellulae or forked to varying degrees; in some cases more strongly anastomosing, veined types of hymenophore can be observed. The colour of the hymenophore is typically very pale, sometimes nearly white when young, but then develops pale yellowish tinges with age, which distinguishes this species from C. albidolutescens, another very similar, usually somewhat paler chanterelle with intensively yellowing context.
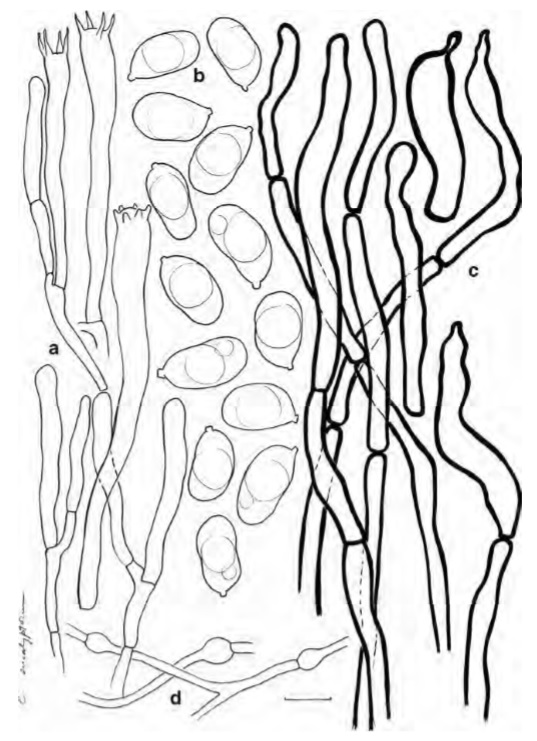
Cantharellus eucalyptorum (holotype) a Spores. b Basidia, basidiola and subhymenial cells c Hyphal extremities of the pileipellis. Scalebars=10μm,butonly5μm for spores. Drawings B. Buyck