Cantharellus luteostipitatus Buyck, Randrianjohany & V. Hofstetter
MycoBank: MB 810671 Facesoffungi number: FoF 00555
Etymology: refers to the partly or entirely bright yellow colouration of the stipe (luteo- meaning yellow and stipus meaning stipe.
Holotype: PC0085575 Pileus mostly 30–50(–60) mm diam., regular or with uneven surface when young and with sunken center from the beginning, the margin strongly inrolled when young and remaining so for a long time, only in older specimens becoming uplifted or revolute at the very extreme border only, sometimes slightly striate over a distance of max. 10 mm, hygrophanous, when wet smooth and with velvety or even distinctly greasy aspect, under a hand lens having almost a frozen aspect, on drying becoming cottony-tomentose toward the center, more rarely fragmenting in larger, appressed squamules, colours varying between reddish pink and yellowish orange tints, often unevenly distributed on the surface and fading rapidly on drying. Hymenophore decurrent, of well-developed gill folds, 2–4(–6) mm high, very variable in colour, starting out as very pale cream or nearly whitish, but sometimes rapidly developing yellowish orange tints, particularly toward the cap margin, gills predominantly unequal, forkings abundant only close to the very cap margin, varying from completely smooth to heavily anastomosed in between gills. Stipe mostly shorter than the cap diam., 8–17 mm diam., subcylindrical or slightly narrowing upwards, tinged with bright chrome yellow to a diluted yellowish orange over part or all of its surface, except for the extreme base which remains generally pure white, solid. Flesh firm and thick beneath the cap center but then rapidly becoming very thin toward the margin, white but with orange-reddish tinges immediately below the cap surface, sometimes showing distinct bluish tinges when cut, on the surface of stipe and gills yellowing when handled. Taste mild. Smell of apricot. Spore print pale yellowish. Spores ellipsoid, (7.1)7.5–8.04–8.5(8.8)×(4.8)5.2–5.77–6.3(6.7) μm, Q=(1.25)1.31–1.40–1.49 (1.56), smooth, filled with numerous oily inclusions. Basidia 50–60×7–9 μm, subcylindrical to weakly clavulate, (2)4(–5)-spored and with 5–8μm long sterigmata (longer for 2–3 spored basidia); basidiola not particularly irregular in form. Subhymenium pseudoparenchymatic, of large voluminous cells. Cystidia none. Pileipellis a loose (tricho)cutis of ramifying, thin-walled hyphal extremities, measuring mostly 5–10μm diam., terminal cells subcylindrical and undifferentiated or sometimes remarkably short and occasionally more inflated to claviform; incrusting pigments most distinct on the narrowest hyphae of subpellis and trama. Clamp connections absent from all tissues.
Material examined: MADAGASCAR, Province of Toamasina, East Coast, 15 km south of Brickaville, on a private property near Ambila Lemaitso, growing in groups of 3 to 6 individuals under Uapaca littoralis in the deep sandy soils of the littoral forest, 27 June 2011, Buyck & V. Hofstetter 11.042 (PC0085573, paratypus), GenBank TEF: KP033509; Buyck & V. Hofstetter 11.044 (PC0085575, holotypus), GenBank TEF: KP033510; Buyck & V. Hofstetter 11.045 (PC0085576, paratypus), GenBank TEF: KP033511. MADAGASCAR Central Plateau, Ibity, in Uapaca bojeri woodland, 1500–1700 m alt., 27 January 2008, Buyck & V. Hofstetter 08.210 (PC 0085130).
Notes: Madagascar is extremely rich in chanterelles (Buyck 2012, 2014; Buyck et al. 2014; Buyck and Randrianjohany 2013) with many species showing very close relationships to mainland African taxa. The newly described chanterelle in this paper strongly resembles the type variety of the mainland African Cantharellus platyphyllus Heinemann in the field because of the pinkish red tinges in its cap colour and palecoloured hymenophore of distinct gill folds. The Malagasy C. platyphyllus ssp. bojerienis Eyssart. & Buyck, which is common and abundant in the Uapaca bojeri woodlands on the Central Plateau, has more grayish-yellowish-greenish-orange tints on the cap. Our species differs from both essentially in the bright yellow colouration of its stipe. Nevertheless, there is quite some variation in the general habit and colouration of these taxa. The only feature so far that allowed to separate all the forms of C. platyphyllus from the mainland African C. symoensii Heinemann, a red-capped species that typically develops a deep, egg-yolk yellow hymenophore at maturity, was the form of the spores: shortly ellipsoid to ellipsoid (mean length-width ratio ca 1.4) for C. platyphylllus and its various infraspecific taxa versus narrowly ellipsoid to elongate spores for C. symoensii (mean length-width ratio of ca 1.9– see Eyssartier and Buyck 1999). Our new species, although genetically closer to C. symoensii, has the same spores as C. platyphyllus and therefore makes the identification of the African taxa in subgen. Afrocantharellus Eyssart. & Buyck even more delicate. All specimens of C. luteostipitatus were collected in the deep sandy soils of the littoral forest, but are genetically close to one collection (as C. luteostipitatus cf) made on the rocky, lateritic soils of the seasonally much colder woodlands of the Central highlands near Ibity at 1500–1700 m altitude. The latter collection consists of a single very immature specimen that lacks the yellow stipe and, therefore, looks even more like typical mainland C. platyphyllus. As these ecologically and climatically very different sites harbor mutually exclusive host tree species, it would seem surprising that their associated fungi were less affected. Additional collecting will be needed to solve the status of this Central Highland’s collection. Cantharellus luteostipitatus is the second species of subgenus Afrocantharellus described from Madagascar. Whereas this subgenus was for many years exclusively known from Africa, its distribution has very recently been extended to Malaysia (Buyck et al. 2014) and China (Shao et al. 2014).

Cantharellus luteostipitatus (PC0085576). General habit (photo B. Buyck)

Cantharellus luteostipitatus (holotype) a Spores. b Basidia, basidiola and subhymenial cells. c Hyphal extremities of the pileipellis. Scale bars=10μm, but only 5μm for spores. Drawings B. Buyck
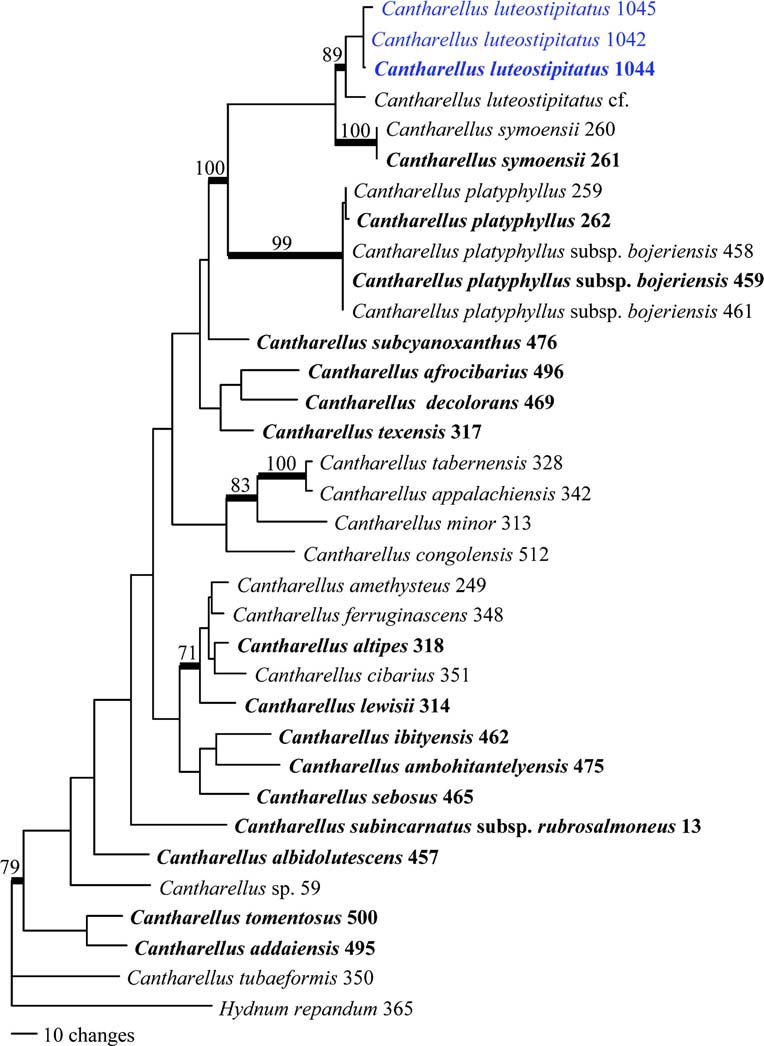
Phylogram generated from Maximum Parsimony analysis based on TEF1 sequence data of Cantharellus. Sequences used in this study have been sampled from Buyck et al. (2014) or newly generated for C. luteostipitatus sp. nov. Branches indicated in bold received significant (≥70 %) bootstrap support. Species for which obtained sequences are based on type material have names in bold
Read more: http://link.springer.com/article/10.1007/s13225-015-0324-y