Amanita emodotrygon Mehmood, Tulloss, K. Das, Iqbal Hosen & R.P. Bhatt
Index Fungorum number: IF552463 Facesoffungi number: FoF 2947
Etymology: emodos (Grk., India) ? trygon (Grk., dove), to indicate possible genetic grouping with North American A. trygonion Tulloss, Pastorino & Kudzma nom. prov. (www.amanitaceae.org/?Amanita%20trygonion)
Holotype: CAL 1338
Pileus 96–132 mm wide, initially convex then campanulate, initially brownish-grey to greyish-brown (7E2–3) and finally olive (1E4–4E7) smooth, dry, shiny. Pileus context 8–10 mm thick, first thinning slowly then rapidly toward margin, membranous at margin, off-white, unchanging when bruised or exposed. Margin with distinct regular veins or striations and corresponding grooves (up to 36 mm wide few veins forked more toward margin, nonappendiculate, whitish to greyish in between the grooves. Universal veil on pileus absent Lamellae free, crowded
(11–13 per 10 mm at margin), 6 mm broad, initially white (1A1) fading pale yellow with age. Lamellulae truncate, of 3–4 lengths, unevenly distributed. Stipe 150–183 x 16–19 mm, equal or slightly tapering upward initially off-white
then greenish-grey to greyish-green (1C2–3), with olive brown tinge toward the base. Context off-white, hollow, stuffed in button stage. Partial veil absent. Universal veil at stipe base saccate, 33–41 x 19–29 mm, white to off-white (1A1–2A1) with many olive-brown spots on exterior surface when age, bilimbate, membranous, thick at bottom thin toward margin. Spore print cream. Odor indistinct. Taste not recorded. Basidiospores [60/3/1] (8.5–)9.8–12
(–13.5) x (8–)9.5–11(–13) um (L = 10–11.5 um; L0 = 10.9 um; W = 9.5–10.5 um; W0 = 10.2 um; Q = (1–) 1.05–1.12(–1.17); Q = 1.05–1.11; Q0 = 1.08), globose to subglobose, rarely broadly ellipsoid, inamyloid, hyaline, colourless, thin walled, with uniguttulate contents, apiculus lateral to sublateral to 1.5 x 1.2 um. Basidia (46–)52–65 (–70) x (14–)14.5–16(–17) um, 2- to 4-spored, thin-walled, colorless; sterigmata 4–6 x 1.5–2 um. Clamp connections not observed at the base of Basidia. Subhymenium wst-near = 35–65 um; wst-far = 50–75 um, Basidia arising mostly from inflated cells to irregular cell (up to 10 x 8 um wide). Lamellae edge sterile; inflated cells; pyriform or clavate 43–48 x 13–22 um, colourless, thin-walled, infrequent; clamp connections not observed. Hymenophoral trama bilateral, divergent; wcs = 70–90 um filamentous undifferentiated hyphae (4–9) um wide; inflated cells 19–27 x 38–45 um; vascular hyphae not observed. Pileipellis 125–190 um thick, in two layers; slightly gelatinized colorless suprapellis (30–60 um thick) of radially arranged filamentous undifferentiated hyphae 1–3 um wide, thin-walled, colourless; nongelatinized subpellis (90–130) um thick); filamentous undifferentiated hyphae 2–7 um wide, subradially and compactly arranged with brown intracellular pigment; vascular hyphae not observed. Pileus trama filamentous, hyphae 2–12 um wide, constricted at septa, slightly thick-walled, hyaline, branching, acrophysalides common up to 135 x 31 um, slightly thickwalled, vascular hyphae up to 17 x 3.5 um. Exterior surface of universal veil (on stipe base) filamentous, undifferentiated hyphae 4–12 um wide, slightly thick walled, septate and frequently branched, colourless, hyaline; globose to subglobose cells up to 102 x 108 um; ellipsoid cells up to 116 x 39 um. Interior surface of universal veil (on stipe base) filamentous, undifferentiated hyphae 4–8 um wide; inflated cells subglobose cells to 45 x 41 um, clavate cells up to 78 x 23 um, infrequent. Stipe context longitudinally acrophysalidic; filamentous hyphae 4–9 um wide, acrophysalidic dominating, (151–190 x 31–42 um); Clamp connections not observed.
Habitat and distribution: Solitary to scattered, under Pinus roxburghii in temperate coniferous forest of western Himalaya.
Specimen examined: India, Uttararakhand, Rudraprayag, Hariyali Devi Forest, 1651 m, N3015.955’ E7903.719’, 27 July 2015, T. Mehmood, TM 15-659 (CAL 1338, holotype). GenBnk number ITS:KX539266.
Notes: The combination of macro- and micromorphological features such as longitudinally acrophysalidic stipe context; bilateral, divergent lamella trama, inamyloid spores, absence of a stipe bulb and absence of Basidia clamps place Amanita emodotrygon in Amanita [subg. Amanita] sect. Vaginatae Sensu Yang (Yang 1997).
In the field this species is distinct from other known species of Amanita sect. Vaginatae by its greyish-brown to olive campanulate pileus, which is deeply grooved towards the margin, crowded white lamellae that becomes pale yellowish with age and occurrence in a coniferous forest under Pinus roxburghii. Micromorphologically, presence of globose to subglobose spores, branched, septate and thick-walled, filamentous hyphae in the universal veil at the stipe base are also quite striking. Based on the key in Tulloss (1994) and (www.amanitaceae.org?stirps% 20Pachyvolvata), there is a group of North American and European taxa that are morphologically similar to A. emodotrygon in having a large, robust basidiome and universal veil-strips Pachyvolvata of Tulloss. The relevant section of the key is largely based on cap colour and spore size and shape. Two of the taxa are described as having an olivaceous tint to the cap. Two are described as having a ring around the inner ends of marginal striations. Hence, gross morphology of this group is similar to that of A. emodotrygon. One of the species has both the cited Pileus characters and is, hence, most similar to the present taxon:
Amanita magnivolvata Aalto (1974) (originally reported from Finland) is segregated from A. emodotrygon by its gray ring on the inner edge of the marginal striations, white Lamellae becoming gray with age and a white exterior surface of the universal veil, developing small, rusty, yellow spots. It occurs in mixed forest including Alnus, Betula, and Picea in Scandinavia and has been reported from Italy (Liguria). Its spores are subglobose to broadly ellipsoid and have comparatively higher Q values than occur in the present taxon-Q = (1.03–) 1.06–1.36 (–1.52); Q = 1.18. (www.amanitaceae.org?Amanita% 20magnivolvata)
Our nrLSU-tree clearly indicates the genetic dissimilarities of Amanita emodotrygon from three taxa. BLAST search indicates relative closeness (98% identity with 98% query coverage) of Amanita emodotrygon to Amanita trygonion (GenBank numbers KU186810); (94% identity with 94% query coverage) of Amanita emodotrygon with Amanita penetratrix (GenBank numbers KU186834, KU186833, KU186832, KU186831); (93% identity to 93% query coverage) of Amanita emodotrygon to Amanita myrmeciae (GenBank numbers KU852505 and KU186806).
The relatively small number of taxa that have been sampled for nrLSU in section Vaginatae and the great difficulty in dealing with the genus by morphological means alone create an unusual situation in the present case, i.e., all of the sequences for which close BLAST comparisons exist in the case of A. emodotrygon are taxa described provisionally. Hence, we can say, at the time of writing, the present taxon is quite distinct from all published species of sect. Vaginatae for which sequence data exists.
Amanita trygonion Tulloss, Pastorino & Kudzma, nom. prov. is a small gray species of the sandy coastal plain in southeastern Texas, USA, where it occurs in Pinus dominated mixed forest and has subglobose to broadly ellipsoid spores with higher Q values than those of the present taxon-Q = (1.05–) 1.10–1.33 (–1.40); Q = 1.17–1.20; Q’ = 1.19. (www.amanitaceae.org?Amanita% 20trygonion)
Amanita penetratrix Tulloss & Kudzma, nom. prov. is a large species with a hard-fleshed, prominent umbo in the center of a gray to gray-brown to white Pileus. It is known from a few sites in the northeastern USA. It occurs in mixed forest including Pinus strobus, Betula, Fagus, and Quercus and has spores that are dominantly subglobose to broadly ellipsoid with higher Q values than those of the present taxon-Q = (1.04–) 1.05–1.25 (–1.34); Q = 1.09–1.16; Q’ = 1.12. The primordium of this mushroom develops deep in forest soil. The unusual umbo allows penetration of up to 154 mm of soil dense with roots and is reminiscent of the perforatorium of species of Termitomyces. (www.amanitaceae.org?Amanita% 20penetratrix)
Amanita myrmeciae Tulloss, Kudzma & Albertella, nom. prov. is known only from association with eucalypts in the Blue Mountains of New South Wales, Australia. This species is unique among those discussed here by being a significant food source for ants of the genus Myrmecia. The species is known from one specimen with a light brown to brown Pileus 62 mm wide and has subglobose spores with slightly higher Q values than those of the present taxon—Q = 1.06–1.18; Q 1.10. (www.amanitaceae.org?Amanita% 20myrmeciae)
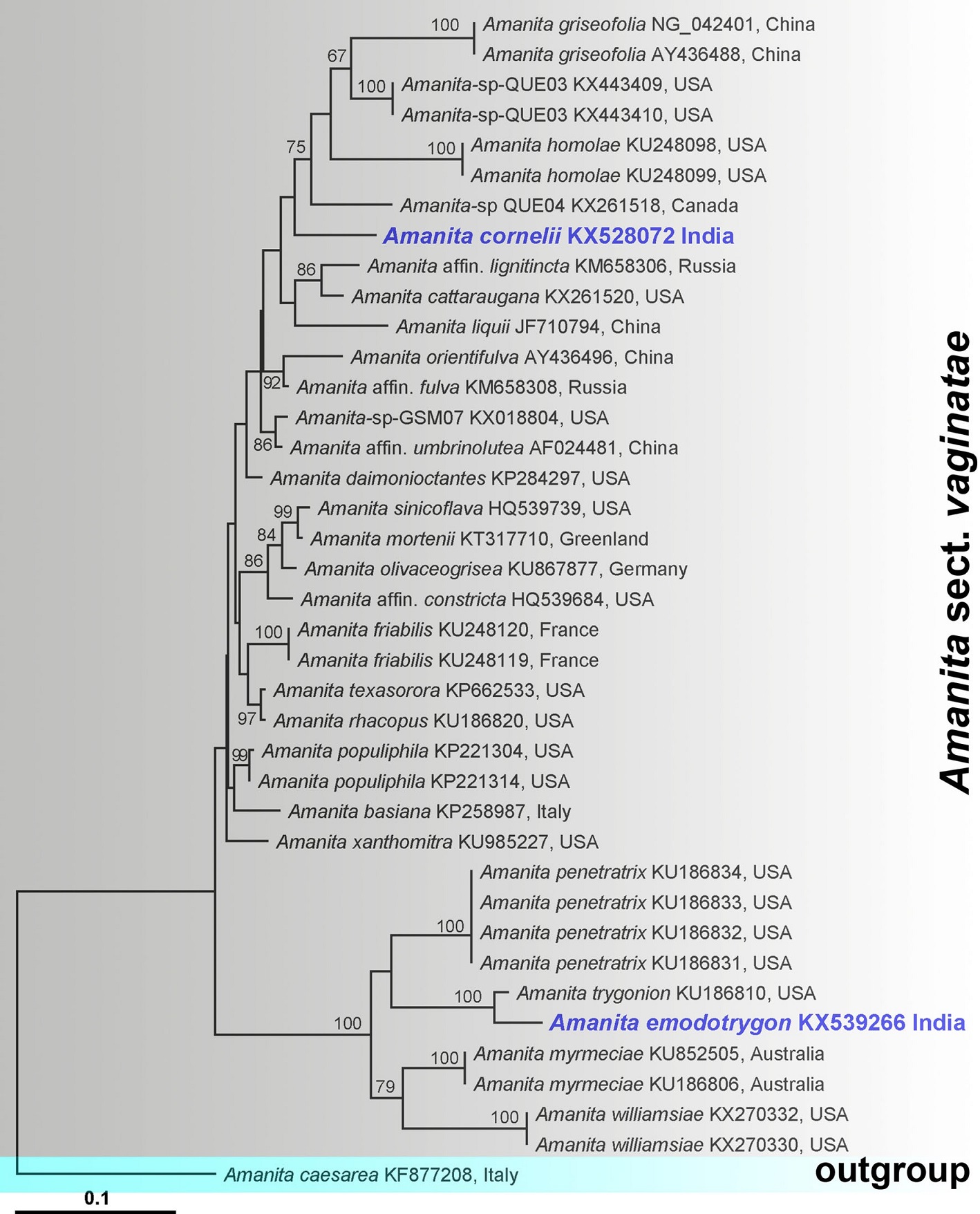
RAxML tree and putative relationships of Amanita subg. Amanita sect. Vaginatae inferred from analysis of LSU sequence data. Bootstrap support values ([50%) obtained from maximum likelihood analysis are shown above or below the branches at the nodes. The new taxa are highlighted in bold and blue font in the tree. GenBank numbers accession numbers are provided after each species name and followed by country of origin. Amanita caesareae (Scop.) Pers. served as the outgroup taxon. The tree is artificial in that it embodies two somewhat distant clusters of taxa associated with the two new taxa by morphology and/or BLAST searches. This results in a compact single figure and, also, in low support for nodes near the tree’s root

Amanita emodotrygon (CAL 1338, holotype). a-c Fresh basidiomata in the field. d Basidia and basidiole. e Elements of universal veil from stipe base. f Basidiospores. Scale bars: b = 100 mm, d-f = 50 µm.
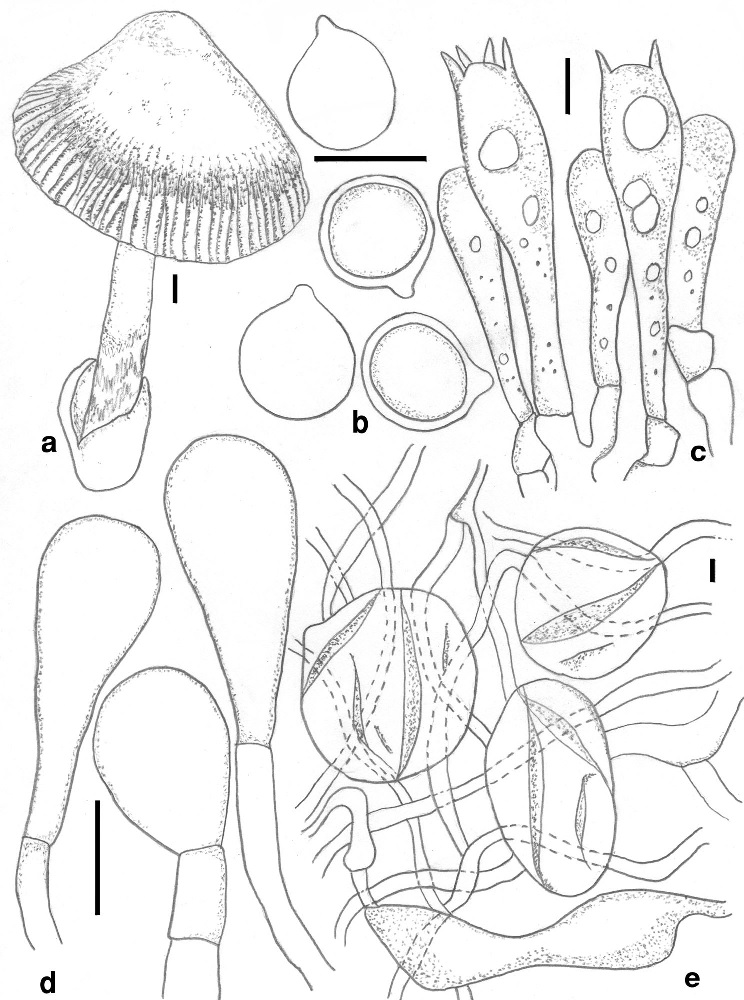
Amanita emodotrygon (CAL 1338, holotype). a Basidiomata. b Basidiospores. c Basidia and element of subhymenium. d Lamellae edges cells. e Elements of universal veil at stipe base. Scale bars: a = 10 mm, b-e = 10 µm.