Psilocybe ruiliensis T. Ma, X.F. Ling & K.D. Hyde, sp. nov., Index Fungorum number: IF 552166, (Figs. 1, 2)
Etymology: the species epithet ‘ruiliensis’ refers to the location Rili where the type collections were found.
Holotype: CHINA, Yunnan Province: Dehong Dai-Jingpo Autonomous Prefecture, Ruili, Longchuan County, on grassland near a reservoir, E097°48´03.6”, N24°15´48.9”, 940m, 24 September 2011, Tao Ma, Xiao-Fei Ling RLC005 (IFRD 415241).
Diagnosis: the new species is characteristic by growing on grassland in the southern subtropical region, basidiomata are somewhat bluish when touched or bruised, having bright reddish orange-brown to yellowish brown and hemisphaeric or subhemisphaeric pileus, the pileus with or without umbo, ellipsoid to subhexagonal basidiospores of medium size 9–11 × (5.5–) 6–7.5 × 5.5–7 μm, slender to stout and fusiform to clavate cheilocystidia, and the form of pleurocystidia similar to the stout pleurocystidia.
Original description: Pileus 10–20 mm diameter, at first irregularly hemisphaeric, conic to subcampanulate, bright reddish orange-brown (5B8), then hemisphaeric, hemisphaeric-conic, hemispheric-convex to almost plane, with or without umbo or small acutely papillate at the disk, yellowish orange-brown (5B6) or dirty yellowish brown when moist, and often with reddish tinge, straw yellow or brownish when dry, frequently whitish at edge; surface smooth, hygrophanous and more or less translucently striate when moist; margin often decorated with somewhat fugacious white to bluing veil remnants, and with cortinate white veil and sometimes small scales in young stages; context yellowish (1A2), sometimes turning bluish when bruised in matured stages, watery brown when wet. Lamellae adnate to subsinuate or adnexed, 1.7–4.2 mm high, subdistant to close, yellowish or beige (4A2) when young, then becoming brownish buff, earth yellow, yellowish brown to chocolate brown (6E7) with gray-purple or purple tinge; edges serrulate, slightly wavy, and remaining whitish and sometimes slightly blackish. Stipe 27–62 mm × 1.5–3.5 mm, central, occasionally slightly eccentric, equal to slightly bulbous at the base, straight to flexuous, yellow-white to brownish, somewhat bluish when touched or bruised; surface covered with appressed whitish pruinose-fibrils; base of stipe with white mycelium; context hollow, fibrous or soft, whitish to brownish, bluish when bruised; annulus absent. Odour null or with somewhat fresh grassy smell.
Microscopic characteristics: Pileipellis an ixocutis, 15–60μm thick, made up of creeping, hyaline and colourless, subregular to interwoven, 2–5 μm wide filamentous to slender tubular hyphae, sometimes including dark yellow and occasionally blue intracellular pigments, wall smooth or with finely incrusted pigments; subpileipellis yellowish brown to brown in KOH, composed of colourless to dark yellowish or brownish, tubular to inflated, 5–17 μm wide hyphae, and the wall often rough for incrusted pigments. Subhymenium subcellular, hyaline, composed of irregular vesiculose to polygonal or subglobose cells. Hymenophoral trama dark yellowish or brownish yellow, regular, with cylindrical hyphae 3–19 μm diameter, hyaline, colourless to yellowish, thin-walled to slightly thick-walled (≤1.0 μm), and the wall smooth to with somewhat dark incrusted pigments. Basidia (18.5–) 21–29.5 (–32) × (7–) 8.5–11 (–12) μm, hyaline and colourless, nearly cylindric to clavate, often narrowed in lower half and constricted in the middle, 4-spored, sometimes 2-spored; sterigmata (2–) 3–5.5 (–7) μm long. Basidiospores (345/10/5) (7.5–) 9–11 (–12.5) × (5.5–) 6–7.5 (–8) × 5.5–7 (–7.5) μm, ellipsoid, ovoid, subrhomboid to subhexagonal in face view, Q = (1.2–) 1.3–1.6 (–1.8), Q = 1.49±0.10; ellipsoid or subellipsoid in side view, Q = (1.4–) 1.5–1.9 (–2), Q = 1.64±0.12, sometimes containing 1–2 oil drops, brown with purple tinge in water, dark yellow in KOH, purplish brown in deposit; wall smooth, slightly thick (0.5–1 μm), complex, with distinct 1–1.5 μm wide apical germ pore. Cheilocystidia (14–) 16–26 (–30) × 3.5–8.5 (–9.5) μm, slender to stout, hyaline, fusiform, fusiform-lageniform to fusiform-clavate or clavate, sometimes sublageniform or broadly lageniform, and the stout similar to pleurocystidia, but bigger, often with a 1.5–9.5 × 1.5–2.5 μm rostrum or neck, occasionally forked, the top or apex frequently seems wall thickened or contain some matter. Pleurocystidia scattered, 13.5–22 (–24) × 5.5–9 μm, thin-walled, hyaline, fusiform to fusiform-clavate or clavate, sometimes sublageniform or ventricose, apex often protruded or extended into a 0.5–4.5 (–6) × 1.5–3.5 μm rostrum or sometimes neck, the top or apex frequently with a thickened wall or containing some matter. Caulocystidia relatively rare, scarce in young individuals, (16–) 22–70 (–90) × (4.5–) 5.5–15.5 (–24.5) μm, often clustered at the upper part of the stipe, thin-walled, hyaline, elongated lageniform, fusiform-lageniform, ventricose, clavate, obclavate or nearly cylindric, occasionally ping-pong racket shape, often with an elongated (2–) 5–26.5 (–44.5) × 1.5–3.5 μm neck, the top or apex frequently with a thickened wall or containing some matter. Clamp connections common in all parts of the basidioma.
Distribution: Rili, Yunnan Province, southwest China.
Habitat: Growing solitary to scattered and gregarious on grassland near a reservoir where cattle and horses have grazed in early autumn.
Material examined: CHINA, Yunnan Province: Dehong Dai-Jingpo Autonomous Prefecture, Ruili, Longchuan County, on grassland near a reservoir, E097°48´03.6”, N24°15´48.9”, 940m, 24 September 2011, Tao Ma, Xiao-Fei Ling RLC005 (IFRD 415241, holotype), RLC004 (IFRD 415240, paratype), RLC006 (IFRD 415242, paratype), RLC007 (IFRD 415243, paratype) and RLC008 (IFRD 415244, paratype).
Note: Psilocybe ruiliensis was only found growing on grassland in the southern subtropical region (close to tropical) of western Yunnan at a relatively low altitude. It can be distinguished by its bright reddish orange-brown to yellowish brown and hemisphaeric or subhemisphaeric pileus, ellipsoid to subhexagonal basidiospores with medium size in face view, slender to stout cheilocystidia, and the presence of pleurocystidia. Phylogenetically, P. ruiliensis forms a monophyletic clade with high BS support (100%) and pp value (1.00) and groups with P. mexicana, P. tampanensis and P. samuiensis (sect. Mexicana) and P. caerulescens (sect. Cordispora) sensu Guzmán (1995) (Fig. 3). In this group, P. ruiliensis and P. samuiensis Guzmán, Bandala & J.W.Allen cluster together.
These taxa are similar in some features of their basidiomata, habitat and basidiospores, but P. samuiensis differs in having a smaller and convex to conic-convex to campanulate pileus and relatively slender stem. Microscopically, the slightly larger hexagonal and subrhomboid basidiospores (9.6–12.0 × 6.4–8.4 μm), the ventricose-lageniform cheilocystidia and pleurocystidia, and the relatively slender pleurocystidia and basidia can easily separate P. samuiensis from P. ruiliensis (Guzmán et al. 1993, Ramírez-Cruz et al. 2013). Morphologically, P. ruiliensis is related to stirps Mexicana according to Singer (1986) because of the medium sized spores, mycenoid habit and growing in open places on soil. It is also related to Sect. Mexicana sensu Guzmán because of the size of spores, the mycenoid habit and the subtropical habitat, although the shape of its basidiospores is partly subrhomboid. In sect. Mexicana sensu Guzmán, besides P. samuiensis mentioned above, P. ruiliensis bears some superficial resemblance to P. mexicana. Both species grow on subtropical grasslands, but the different shape of fruit body, the absence of pleurocystidia, and the subrhomboid spores of P. mexicana easily distinguish them (Singer and Smith 1958, Guzmán 1983). In many aspects, P. ruiliensis is also similar to P. gallaeciae Guzmán & M.L. Castro and P. galindii Guzmán, but the latter two species have pseudorhiza. In addition, P. gallaeciae has a different coloured pileus, slightly broader and thicker-walled spores, and lageniform pleurocystidia and cheilocystidia. Psilocybe galindii has relatively slender basidiomata, smaller basidia, and slightly larger and subrhomboid thicker-walled basidiospores, as well as grows at 1700–1800 m elevation (Guzmán 1983, Guzmán & Castro 2003, Ramírez-Cruz et al. 2013).

Fig. 1. Basidiomata of Psilocybe species. A–G. Psilocybe keralensis. A, E: IFRD415173, B: IFRD415174, C–D: IFRD414040, F–G: IFRD414213; H, L: Psilocybe aff. fasciata, IFRD415224; I–K, M–N. Psilocybe ruiliensis, I–J: IFRD415243, K, N: holotype, IFRD415241, M: IFRD415240.
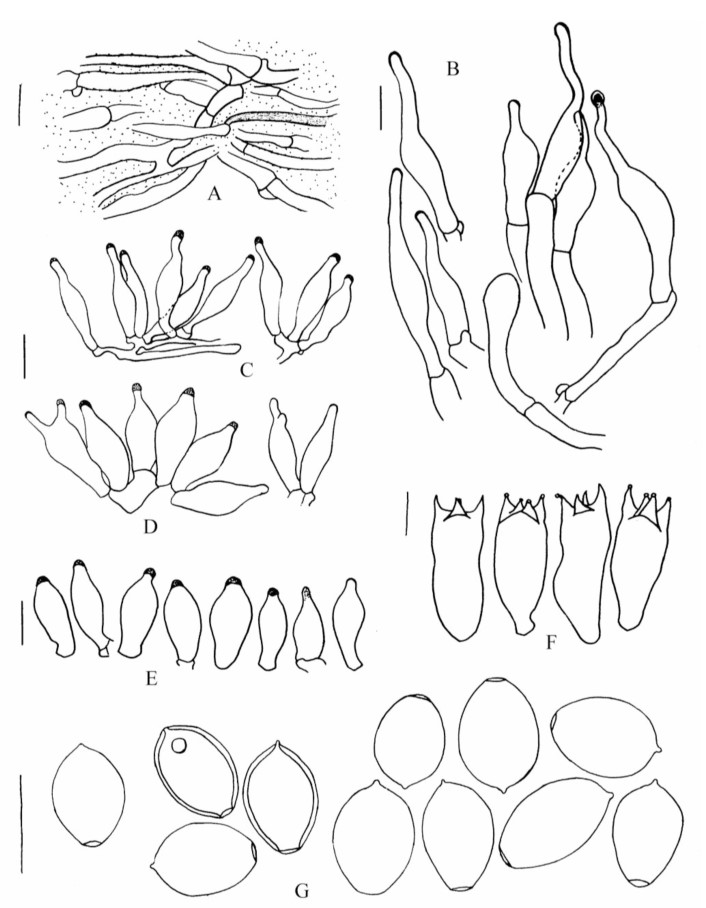
Fig. 2. Psilocybe ruiliensis A–C, E–G. (holotype, IFRD415241). D. (IFRD415240), A: Pileipellis, B: Caulocystidia, C–D: Cheilocystidia, E: Pleurocystidia, F: Basidia, G: Basidiospores. Scal bar=10 μm.
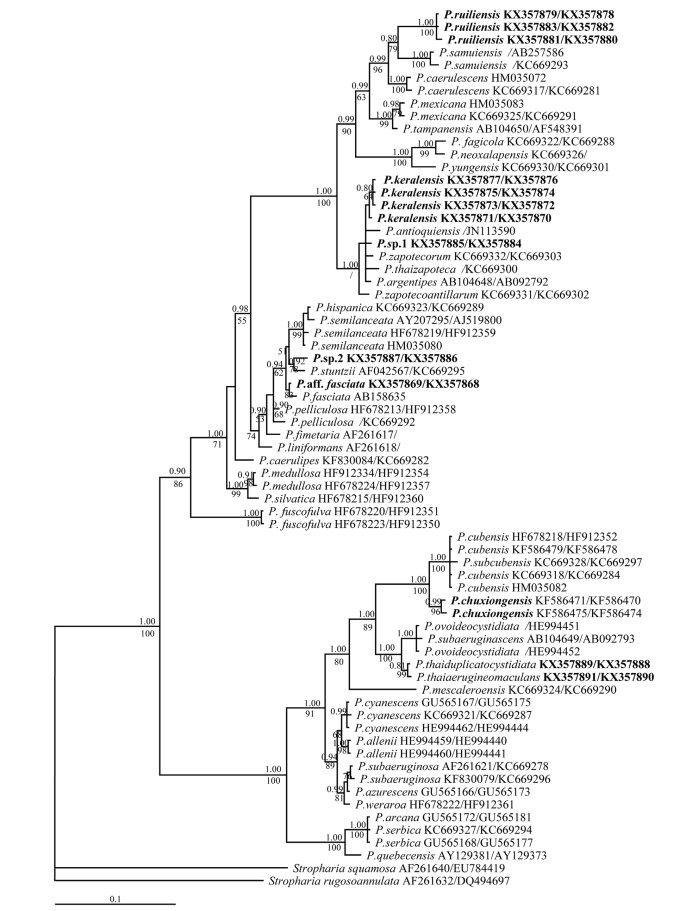
Fig. 3. Phylogram inferred from analyses of a combined LSU and ITS dataset using MrBayes. Values above and below branches are posterior probabilities (pp≥0.80) and RAxML bootstrap (BS≥50%) values. For node with little difference between RAxML and BI analyses, only the pp value is displayed and BS value is not shown but indicated by ‘/’. The terminal taxa represent individual specimens with GenBank accession numbers. nrLSU and ITS numbers are indicated as LSU/ITS and the sequences that are lacking are not shown, and only one number is represented the sequence that is including nrLSU and ITS data. Species from Yunnan province and new sequences generated in this study are in bold.