Cordana terrestris (Timonin) Hern.-Restr., Gené & Guarro, in Hernández-Restrepo et al., Mycologia 106, 729 (2014)
Index Fungorum number: IF 807979; Facesoffungi number: FoF 05475; Fig. 1
Hyphae septate, branched, smooth, subhyaline to hyaline, becoming rough and brown when mature. Asexual morph: Conidiophores 25–73 × 1.5–3.5 μm (x̅ = 44 × 2 μm, n = 20), micronematous, mononematous, solitary, erect, unbranched or branched, septate, straight or flexuous, cylindrical, hyaline to sub-hyaline. Conidiogenous cells holoblastic, polyblastic, terminal, discrete, hyaline. Conidia 4–7 × 2.5–4.5 μm (x̅ = 5 × 3.5 μm, n = 20), terminal, mostly oblong, sometimes ellipsoid or sub-cylindrical, guttulate, sometimes mid-region slightly constricted, hyaline, smooth-walled. Sexual morph: Undetermined.
Culture characteristics – Colonies on PDA become 30 mm in diam. after 14 days at 25 ℃, flat, effuse, dark gray to black-brown, undulate margined, reverse grayish brown at the center, become dark brown to brown at the margin.
Material examined – Thailand, Chiang Rai Province, May Yao, from forest soil, 20.0478N 99.7619 E, 863m, 23 September 2019, W.A.E. Yasanthika, living culture, MFLUCC 23-0022.
GenBank accession numbers – ITS: OQ690712, LSU: OQ690715.
Known hosts (based on molecular data) – Soil, Cecropia leaves, decaying, submerged wood (Timonin 1940, Fernández & Huhndorf 2004, Luo et al. 2019, this study).
Known distribution (based on molecular data) – China, Brazil, Canada, Cuba, Democratic Republic of Congo (Zaire), Jamaica, Japan, India, New Zealand, Panama, Poland, Puerto Rico, the USA (Georgia, Iowa) (Fernández & Huhndorf 2004, Luo et al. 2019), Thailand (this study).
Notes – Cordana terrestris is widespread and has been identified in various aquatic and terrestrial habitats (Hyde et al. 2020a). This species was previously recorded from the soil in Georgia (Timonin 1940). Spicularia terrestris and Pseudobotrytis fusca were synonymized as C. terrestris by Hernández-Restrepo et al. (2014). Our collection (MFLUCC 23-0022) phylogenetically clustered with other C. terrestris isolates with 92% maximum likelihood bootstrap support and 1.00 Bayesian posterior probability in LSU-ITS multigene phylogenetic analyses (Fig. 2). The asexual morph of our isolate (Fig. 1) is similar to C. terrestris in having terminal, mostly oblong, ellipsoidal or sub-cylindrical, guttulate conidia (Fernández & Huhndorf 2004, Hernández-Restrepo et al. 2014, Luo et al. 2019). Conidial septation and macronematous conidiophores are absent in our isolate in contrast to C. terrestris described in Fernández & Huhndorf (2004) and Luo et al. (2019). These morphological differences can result from environmental adaptations (Francisco et al. 2019). Based on the biphasic approach, we report our collection (MFLUCC 23-0022) of C. terrestris as the first record on Thailand soil.

Figure 1 — Cordana terrestris (MFLUCC 23-0022, a new habitat record). a Colony from above (on PDA). b Colony from below (on PDA). c Conidial attachments. d Immature hyphae. e Mature, pigmented hyphae. f–j Conidiogenesis on the conidiophores. k–o Conidia. Scale bars: d = 20 μm, e = 15 μm, f–i = 10 μm, j, k–o = 5 μm.
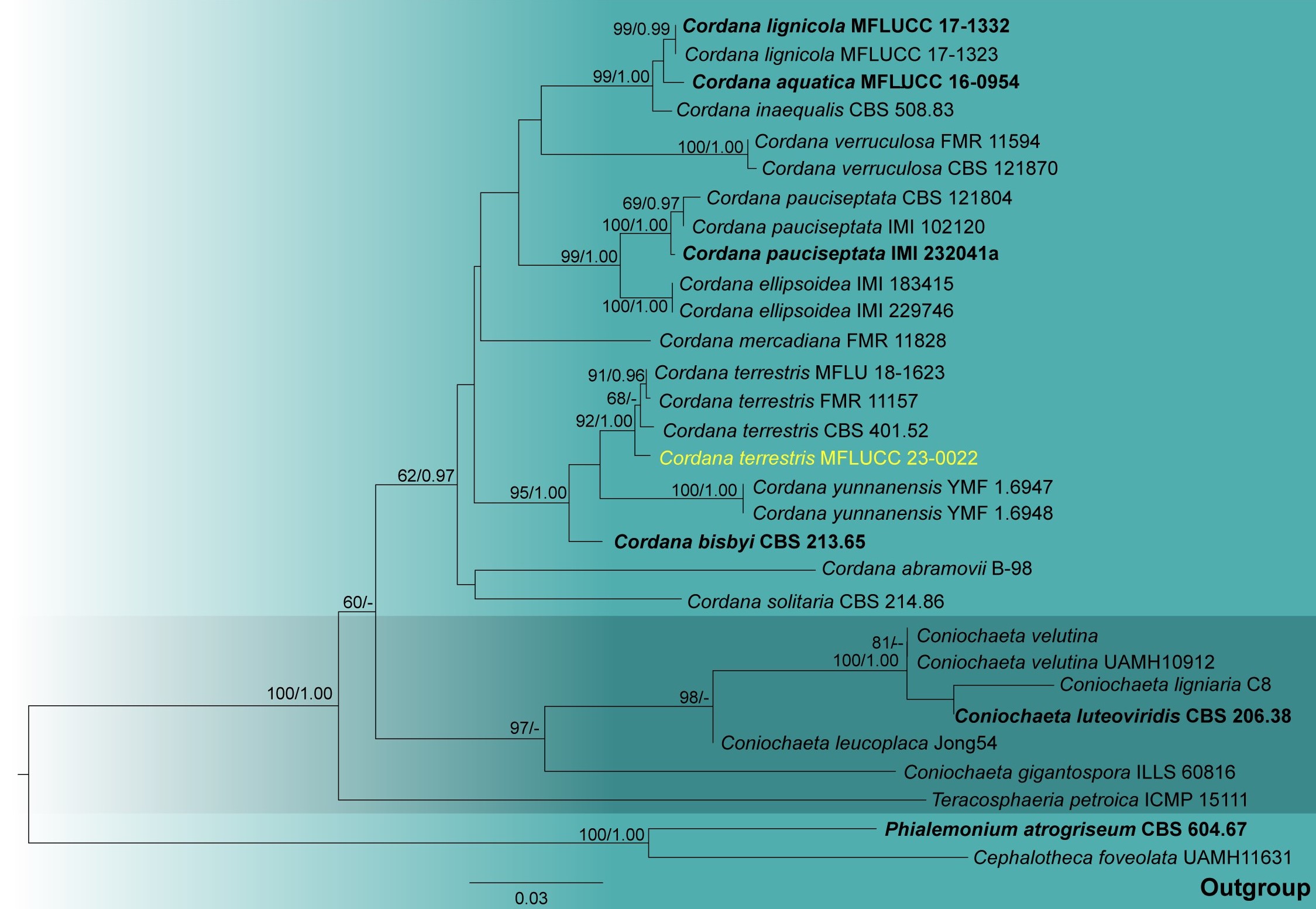
Figure 2 — Phylogram generated from maximum likelihood analysis based on the combined ITS and LSU sequence data of Coniochaetales taxa. Twenty-nine strains are included in the combined analyses, which comprised 1452 characters (595 characters for ITS and 857 characters for LSU) after aligned. Tree topology of the maximum likelihood analysis is similar to the Bayesian analysis. The best RAxML tree with a final likelihood value of -5536.915184 is presented. The matrix had 427 distinct alignment patterns, with 28.19% undetermined characters or gaps. Evolutionary models applied for ITS and LSU are SYM+I+G and GTR+I+G models, respectively. Bootstrap support values for ML equal to or greater than 60% and Bayesian posterior probabilities equal to or greater than 0.95 are given near nodes, respectively. The tree is rooted with Cephalotheca foveolata (UAMH11631), Phialemonium atrogriseum (CBS 604.67) and Teracosphaeria petroica (ICMP 15111). Ex-type strains are in bold. The newly generated sequences are indicated in yellow.