Pleurotus giganteus (Berk.) Karun. & K.D. Hyde in Karunarathna, Mycotaxon 118: 62 (2011), Index Fungorum number: IF 561087, Figs 1–2
≡ Lentinus giganteus Berk., Lond. Journ. Bot. 6: 493[bis], pl.17/18 f.2 (1847)
≡ Pocillaria gigantea (Berk.) Kuntze, Revis. Gen. Pl. 2: 866 (1891)
≡ Velolentinus giganteus (Berk.) Overeem, Bull. Jard. Bot. Buitenz, 3 sér., 9: 12 (1927)
≡ Panus giganteus (Berk.) Corner, Beih. Nova Hedwigia 69: 69 (1981)
= Lentinus maculatus Berk., London Journal of Botany 6: 494, pl. 19/2 (1847)
= Lentinus obnubilus Berk., London Journal of Botany 6: 495, pl. 19/3 (1847)
= Lentinus velatus Berk. & Broome, Botanical Journal of the Linnean Society 14: 42 (1875)
= Lentinus radicans Berk. & Broome, Botanical Journal of the Linnean Society 14: 44 (1875)
= Pocillaria obnubilis (Berk.) Kuntze (1891)
Pilei 40–310 mm in diam., convex to applanate dark brown (7F4) towards margin, reddish grey in the margin (12D3), fibrillose-scaly at the centre, at first uniformly dark at the surface, fuscous brown and black, then fading with age to yellowish brown (E9), with a darker centre, remaining dark, dry, disrupted into small, indefinite, radial, innate squamules, overlaid by scanty, pale grey or blackish, verrucose-floccose, with concentrically arranged remnants of the veil; margin strongly involute, thin. Lamellae moderately crowded with lamellulae of five to six lengths, decurrent, 3–4 mm broad, white to cream (3A1); edge entire, pale ochraceous or yellowish brown (E9). Stipe up to 40–200 mm long, 6–10 mm broad at the apex, 9–15 mm at the base, fusiform, with radicating base (50–70 mm long), solid, surface concolorous with the pileus, paler at the apex, finely tomentose with indefinite zones of paler velar remnants, in the early stages; veil thin, floccose, pale to dark brown (6F5), soon reduced to floccose remnants but never forming an annulus on the stipe. Context 4–10 mm thick at the disk, submembranous at the pileal margin, white in pileus and stipe, fleshy-spongy, consisting of a dimitic hyphal system with skeletal hyphae. Generative hyphae (Fig. 2e) 3–6 μm in diameter, inflating with a thick or slightly thickened wall, more or less radially parallel, but frequently branching and with large clamp connections. Skeletal hyphae (Fig. 2d) 5–8 μm in diameter, hyaline of intercalary or terminal origin, becoming very thick-walled with a narrow lumen, tending to taper apically, occasionally with a limited lateral branch. Basidiospores (Fig. 2a) 6–8.5 (–9) × 4.4–5.3 μm [n =60, Lm=7.65 μm, Wm=5.02 μm, Q=1.36–1.69, Qm=1.51], broadly ellipsoid to ellipsoid, white in mass, smooth, with one large oil drop or multiguttulate, inamyloid, thin-walled. The large basidiospores are not cylindrical but broadly ellipsoid, although the largest basidiospores become oblong ellipsoid. Basidia (Fig. 2c) 28–50 × 8–9 μm, elongate, clavate, bearing 4 sterigmata. Lamella edge sterile with a broad layer of Cheilocystidia (Fig. 2b) 23–36 × 6–10 μm, more or less lecythiform, with a ventricose base and a small capitellum (2–4 μm), subtended by a narrow neck, hyaline, thin-walled.
Habitat and distribution:—on buried rotten wood in rain forest, solitary. Widely distributed in Australia, China, Malay Peninsula, Sabah, Sri Lanka, Vietnam (Pegler 1983), and Thailand (Karunarathna et al. 2011a, b).
Specimens examined: China, Yunnan Province, Xishuangbanna, Mengsong Village; UTM-N2379744.485281, UTM-E653217.753714; alt. 1600m; 27 June 2014; on ground; Jie Chen (MFLU 14-0740); 15 June 2012; Samantha C. Karunarathna (MFLU 14-0741); Jiangxi Province; Fuzhou City; 6 February 2015; from JingShan mushroom growing company; Jinshan Fang (ZRL-2015001, HMAS-253762); SRI LANKA, Central Prov., Kandy Distr., Deliwala village, 7°14′43.55″N 80°33′51.40″E, elevation 1050 m, rainforest dominated by Swietenia spp. and Artocarpus heterophyllus, 5 June 2009; on ground; Samantha C. Karunarathna (MFLU 11-0018, epitype); THAILAND, Chiang Mai Prov., Mae Taeng Distr., Ban Pha Deng, Mushroom Research Centre, 19°17.123′N 98°44.009′E, elevation 900 m, rainforest dominated by Castanopsis armata, Erythrina sp, and Dipterocarpus sp., 8 July 2008; on ground; Ruilin Zhao (MFLU 08-1370); 21 June 2008; on ground; Samantha C. Karunarathna (MFLU 10-0138); 27 June 2008; on ground; Samantha C. Karunarathna (MFLU 08-1371); 6 July 2008; on ground; Samantha C. Karunarathna (MFLU 08-1382); 22 July 2008; on ground; Samantha C. Karunarathna (MFLU 10-0137); 10 July 2010; on ground; Olivier Raspe (MFLU 10-0153); DoiSuthep-Pui National Park, Sangasabhasri Lane to HuaiKok Ma village, 18°48.62′N 98°54.60′E, elevation 1145 m, rainforest dominated by Castanopsis spp., Lithocarpus polistachyus and other trees, 9 June 2008; on ground; Samantha C. Karunarathna (MFLU 10-0136); Chiang Rai prov., Highway No.110 to Mae Sai, Doi Tung, 20°17′37″N 99°48′56″E, elevation 950 m. 15 July 2009; on ground; Samantha C. Karunarathna (MFLU 10-0140); 15 July 2009; on ground; Samantha C. Karunarathna (MFLU 10-0141); 8 August 2009; on ground; Samantha C. Karunarathna (MFLU 10-0142); 8 August 2009; on ground; Samantha C. Karunarathna (MFLU 10-0143); 16 July 2010, Samantha C. Karunarathna (MFLU 10-0154).

Fig. 1. Basidiocarps of Pleurotus giganteus A, B. Pleurotus giganteus from Mengsong, China (A. MFLU14-0741, B. MFLU14-0740) Scale bars: A, B = 10 cm.
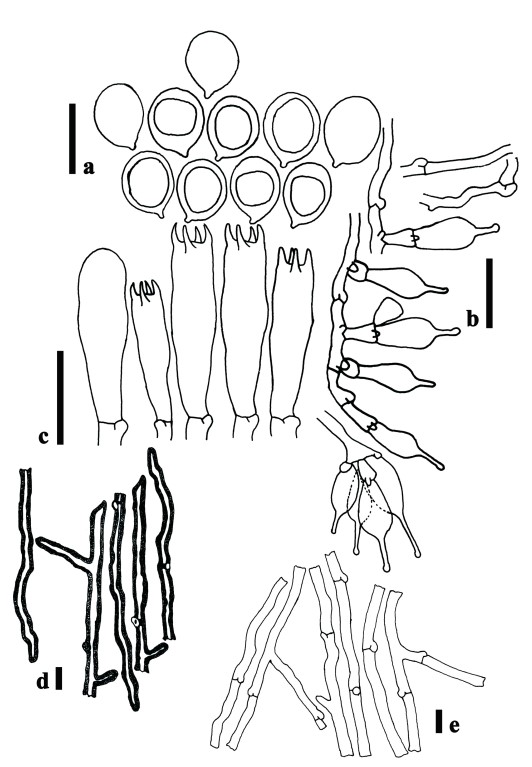
Fig. 2. Pleurotus giganteus (MFLU14-0740/ ZRL-2015001) a. Basidiospores. b. Cheilocystidia. c. Basidia and basidioles. d. Skeletal hyphae. e. Generative hyphae. Scalebars: a–c = 20 μm; d, e = 10 μm.