Humidicutis brunneovinacea R. Garibay-Orijel, sp. nov.
Index Fungorum number: IF 829181; MycoBank number: MB 829181; Facesoffungi number: FoF 05673; Fig. 82
Etymology: Referring to the distinctive colors of the basidiome.
Holotype: MEXU 28225.
Basidiocarp stipitate. Pileus conical, umbonate to campanulate, with age becoming broadly campanulate, planoumbonate or umbilicate with uplifted margin, 10–40(–57) mm diam; lightly lubricous, moist (not viscid) but rapidly drying, hygrophanous, glabrous, finely rugulose when young, radially fbrillose with age; margin incurved and folding when young, with age sometimes becoming crenulate, thin, discolored and often splitting some millimeters; dark vinaceous brown, warm blackish brown or diamine brown (10F6–8) when young, fading towards the margin, in age some basidiocarps stay dark vinaceous, neutral red or bordeaux (12D–E8) while most of them become rose red, pomegranate purple or amaranth purple (12A–C8). Context thin (0.5–1.5 mm), watery, concolorous. Lamellae sinuate or adnate, some with a small decurrent tooth, thin, subdistant with 1–3 series of lamellulae, broad (7–10 mm) margin undulate, white; in age becoming distant, thick, sometimes transvenose to intervenose with red vinaceous margin; whitish to light greyish (10B1) becoming light brown-grey or vinaceous grey (10C–D2) or vinaceous pink (12A–B4) with age. Stipe 40–100×2–10 mm, central, terete, equal or seldom slightly enlarged near the base, straight to curved
in age, moist to dry (not viscid), fragile, glabrous above, base with hirsute short white mycelia, without rhizomorphs; when young apex concolorous with pileus, then becoming spinel red or spinel pink (12A7–6) towards the base, with age it changes frst to pinkish, Persian lilac (12A5), Congo pink or light coral red (9A5–6) and fnally discolors to pale red, shell pink, orange-pink or buf pink (7A2–4). Context hollow, thin (1–1.5 mm), pastel red, coral pink or Congo pink (9A4–5). Odor and taste not distinctive.
Basidiospores broadly ellipsoid to ellipsoid, inequilateral in profile, hilar appendix < 1 µm, hyaline, smooth, non-amyloid, non-dextrinoid, white in deposit, (6–)7–8.5(–9.5) × (4–)5–6(–7) µm ( x̅ = 7.61 × 5.48 µm, n = 20/5), Q = (1.07–)1.25–1.45(–1.6) ( x̅ = 1.39 µm, n = 30/5). Basidia (30–)32–40(–44) × 5–7 µm ( x̅ = 36.7 × 5.86 µm, n = 10/5), Q = 5–7.8 ( x̅ = 6.24 µm, n=10/5), predominantly 4-spored, seldom 3-spored, clavate, hyaline, rare medallion clamp connections and seldom toroidal; sterigmata thin, straight or incurved, (6–)7–10(–11) µm, however in some collections smaller (6–)6–8(–9) µm, these collections have also smaller spores 6–8×4–5 µm. Abundant basidioles often with toruloid clamps and bifurcate base. Hymenial cystidia absent. Pileipellis a weak ixocutis composed of thin-walled, hyaline, non-incrusted, cylindrical, septate, subparallel to slightly interwoven hyphae, (3–)3.5–4.5 µm diam, clamp connections absent. Pileus trama of subparallel to interwoven short, infated to fusoid hyphae 4–8 µm diam. Hymenophoral trama subregular, composed of hyaline, nongelatinous, irregular, hyphal elements (10–)12.5–17.5(–25)×10–12.5(–20) µm, without cylindrical hyphae, clamp connections absent. Stipe tissue monomitic; stipitipellis a cutis composed of parallel or slightly interwoven, thin-walled, hyaline, cylindrical septate hyphae 3.75–5 µm diam, medullary hyphae wider, (15–)17.5–20(–22.5) µm diam, caulocystidia absent, clamp connections absent.
Habit and habitat gregarious and occasionally caespitose, amongst moss in mountain cloud subtropical forest dominated by Cupressus or Quercus spp.
Material examined: MEXICO, Oaxaca, Santa María Huitepec, Cempoaltepetl, Cupressus subtropical cloud forest, 6 November 2018, Garibay-Orijel 2018-1A (MEXU 28225 holotype), Garibay-Orijel 2018-1B (MEXU 28226); Oaxaca, Santa María Huitepec, Quercus spp subtropical cloud forest, 6 November 2018, Garibay-Orijel 2018-2 (MEXU 28227), Garibay-Orijel 2018-3 (MEXU 28228), Garibay-Orijel 2018-4, MEXU 28229).
GenBank numbers: ITS: MK332021, MK332022.
Notes: Phylogenetically, the ITS sequences of Humidicutis brunneovinacea form a distinct terminal lineage together with three undescribed species (Fig. 83). Morphologically, H. brunneovinacea is unique among described species of Humidicutis due its combination of dark brown vinaceous fading to vinaceous and red colors (Fig. 82). There are some rose red undescribed species of Humidicutis in the Caribbean and Central America (Cantrell et al. 2001; Lodge et al. 2014), however these lack dark brown colors and their ITS sequences are too divergent (nucleotide similarity<85%) with H. brunneovinacea to be considered the same species. Microscopically, H. brunneovinacea is distinguished by the very scarce clamp connections in the basidia and the subregular hymenophoral trama composed of irregular hyphal elements without cylindrical hyphae. It is noteworthy that these four species are distributed in the Caribbean and Mesoamerican region of the Neotropics and they share reddish tinges unique in the genus.
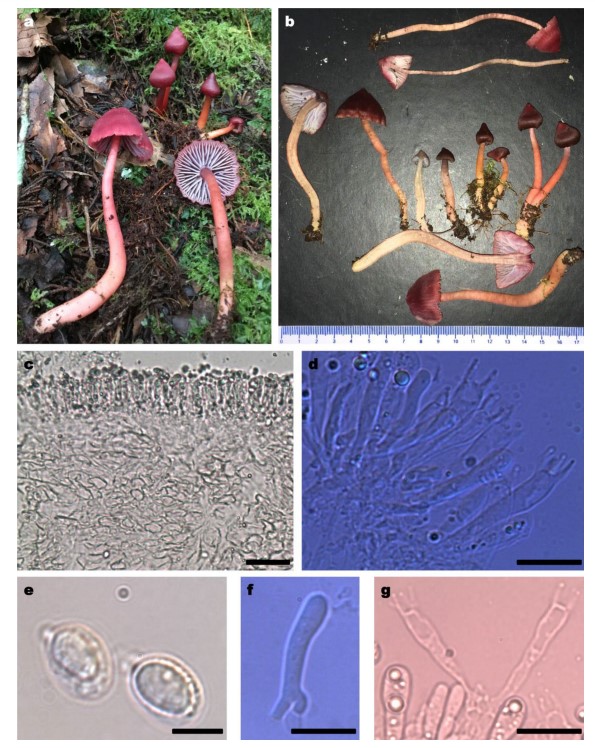
Fig. 82 Humidicutis brunneovinacea (MEXU 28225, holotype). a Basidiomes in its habitat. b Basidiomes development. c Hymenophoral trama. d Hymenia. e Spores. f Basidiole with bifurcated base. g Basidia arising from same toruloid clamp connection. Scale bars: c=40 µm, d, f, g=20 µm, e=5 µm
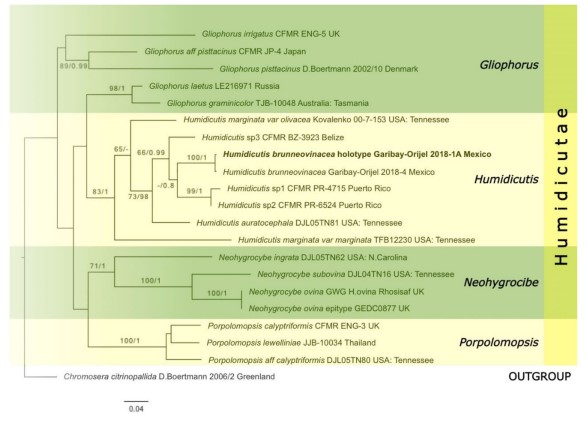
Fig. 83 Maximum likelihood analysis of Humidicutae showing the phylogenetic position of Humidicutis brunneovinacea inferred from ITS and LSU sequences. Bootstrap support values for ML≥60% and Bayesian posterior probabilities≥0.80 are given near nodes respectively. The tree is rooted with Chromosera citrinopallida. The holotype of H. brunneovinacea is in bold