Entoloma calabrum Battistin, Marsico, Vizzini, Vila & Ercole
Index Fungorum number: IF551433 Facesoffungi number: FoF01046
Etymology: the Latin epithet calabrum refers to the Italian region (Calabria) where the species has been first collected.
Holotype: MCVE 28566 Colour notations in the macroscopic descriptions refer to the Munsell Soil-Color Charts (1994). Habit mycenoid. Pileus 19–30 mm broad, at first conicalcampanulate to convex, then applanate with or without a prominent, obtuse umbo, yellow (Mu 10YR 8/6, 8/8), reddish yellow (Mu 7.5YR 6/8, 7/6, 7/8); surface dry, mat, fibrillose, tomentose or pubescent in some specimens, not or hardly hygrophanous; margin at first inflexed than straight, wavy, shortly translucently striate. Lamellae moderately crowded, adnexed to sinuate-emarginate, arcuate, up to 6–7 mm broad, very pale brown (Mu 10YR 8/3, 8/4) with many lamellulae. Stipe 50–60×2–3 mm, central, cylindrical or slightly compressed, equal or tapering at the apex, (sub)concolourous with the pileus, almost completely covered with white fibrils and a white tomentum. Context subconcolourous with pileus; odour not distinctive and taste mild. Spores 6.9–10.4×5.1–8.4 μm, on average 9×6.4 μm, Q=1.16–1.81 (Qm=1.41), almost exclusively heterodiametrical, 5–7 angled in side view. Basidia 35–47 × 11.7–15 μm, clavate, 4-spored, clamped. Sclerobasidia absent. Hymenial cystidia absent. There are some cylindroid, more or less flexuose, thin-walled, sterile hyaline elements scattered among basidia, which scarcely protrude beyond the hymenium. Caulocystidia up to 180 μm long, 5–11.7 μm wide, abundant, cylindrical, straight or curved, flexuose, some coralloid, septate, hyaline. Hymenophoral trama regular in the mediostratum, a bit less in the outer layer, made up of cylindroid or subfusiform hyphae 94–300×3.3–23 μm (n=22; mean: 172×12 μm). Pileipellis a cutis of cylindrical, radially arranged hyphae up to 16.7 μm wide. Pigment very finely encrusting and sometimes also parietal. Clamp-connections present.
Habitat and known distribution: Gregarious on grassy soil. So far known only from southern Italy.
Material examined: ITALY, Calabria, Cosenza, municipality of San Pietro in Guarano, locality Serra Vaccaro, about 39°20′ N 16°24′ E on acid soil in a grassy clearing of a Fagus sylvatica stand near Cytisus scoparius and Astragalus calabrus, 8 October 2010, leg. O. Marsico (MCVE 28566, holotype); same loc., 20 October 2010, leg. O. Marsico (TO AV201010a; same loc., 27 October 201 (TO AV271010a)).
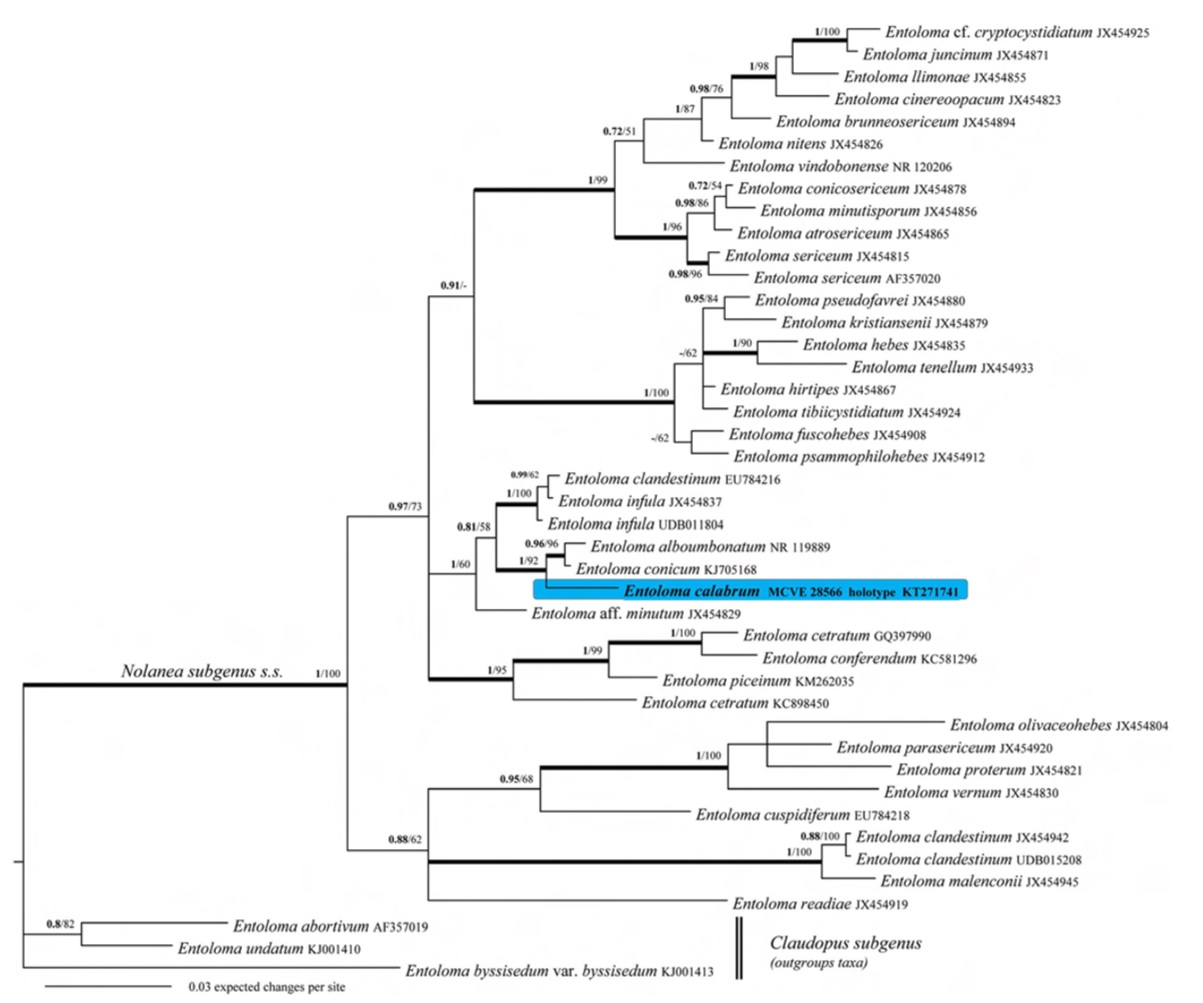
Notes: Both Bayesian and Maximum likelihood analyses produced the same topology; therefore, only the Bayesian tree with both BPP (Bayesian Posterior Probabilities) and MLB (Maximum Likelihood Bootstrap) values. In the ITS sequence analysis the sequence of Entoloma calabrum clustered within the Nolanea major clade, in a clade consisting of E. infula, E. clandestinum, E. conicum, E. alboumbonatum and E. aff. minutum. Macroscopically Entoloma calabrum is a distinct taxon on account of its sometimes zonate, umbonate, strongly fibrillose, almost villose pileus, the brown stipe covered with abundant white fibrils and a white tomentum, the cream or cream-brownish lamellae, and the odourless and mild context. Microscopically it is characterised by the presence of clamp-connections, heterodiametrical spores, pigment mainly finely encrusting but also parietal, absence of hymenial cystidia, and presence of caulocystidia. A combination of morphological features such as size, habit, overall colours and the structure of the hymenophoral trama and pileipellis placed it in the subgenus Nolanea (Fr.: Fr.) Noordel. (typified by Agaricus hirtipes Schumach.) as circumscribed by Noordeloos (1980, 1992, 2004). This placement is supported by the ITS phylogenetic analysis and by the fact that LSU query sequence produced the following matches: 99 % max. identity with Nolanea sericea (accessions GQ289191, AF223171, AY207197 and AF223170) and N. conica (accession AF2613170). According to Noordeloos’ taxonomic arrangement (1992, 2004) and with regard to the five sections included in Nolanea , i.e., Mammosa (Romagn.) Noordel., Cosmeoexonema (Largent & Thiers) Noordel., Endochromonema (Largent & Thiers) Noordel., Fernandae Noordel. and Canosericei Noordel., we think that on a strictly morphological basis E. calabrum should be placed within section Endochromonema subsection Endochromonema (Largent & Thiers) Noordel., especially on account of the heterodiametrical spores and the presence of an intraparietal pigment. Consulting the iconography of Noordeloos’ monograph (1992, 2004), Entoloma calabrum looks vaguely like E. pallescens (P. Karst.) Noordel., E. lanuginosipes Noordel. and E. cetratum (Fr.) M.M. Moser, which belong to section Endochromonema subsection Endochromonema, but the former differs especially by the presence of a subzonate and strongly fibrillose, almost villose pileus and 4-spored basidia. The species gravitating around Entoloma sericeum Quél., some of them recently described (Vila et al. 2013), such as E. minutisporum (Vila & Llimona) J. Carbó et al., E. conicosericeum Vila et al., E. atrosericeum (Kühner) Noordel., E. cinereoopacum (Noordel.) Vila et al., E. vindobonense Noordel. & Hauskn., E. nitens (Velen.) Noordel., E. juncinum (Kühner & Romagn.) Noordel., E. brunneosericeum Noordel. et al., E. llimonae Vila et al., differ from E. calabrum especially in the iso- or subisodiametrical spores and the abundant encrusting epiparietal pigment. Entoloma hebes (Romagn.) Trimbach and closely related taxa like E. hirtipes (Schumach.) M.M. Moser, E. fuscohebes Vila et al., E. psammophilohebes Vila & J. Fernández, E. tenellum (J. Favre) Noordel., E. kristiansenii Noordel., E. tibiicystidiatum Arnolds & Noordel. and E. pseudofavrei Noordel. & Vila can be distinguished from E. calabrum on the basis of the occurrence of distinct cheilocystidia (Noordeloos 2004; Vila et al. 2013). No extra-European known species fits our new taxon (Hesler 1967; Horak 1973, 1976, 1978, 1980, 1982, 2008; Romagnesi and Gilles 1979; Pegler 1983, 1997; Noordeloos 1988a, 2008; Largent 1994; Wölfel and Noordeloos 2001; Manimohan et al. 2006; Gates and Noordeloos 2007; Noordeloos and Gates 2009, 2012; Noordeloos and Hausknecht 2007; Noordeloos and Morozova 2010; Henkel et al. 2014). According to the ITS sequence data analysis, the most closely related species are Entoloma conicum (Sacc.) Hesler and E. infula (Fr.) Noordel. two taxa sometimes with subzonate pileus. E. conicum (Peck) Hesler, described from North America (= E. alboumbonatum Hesler;= E. subquadratum Hesler fide Noordeloos (1988b, 2008), who studied type collections, Horak (1976) and Largent (1994), differs from E. calabrum mainly in having a subfarinaceous odour and taste, isodiametrical, and quadrate to 5-angled spores, Qm=1.2 (Hesler 1967; Noordeloos 1988b). E. infula is distinguished by its subglabrous pileus and stipe surfaces, presence of intracellular and encrusting pigment in the pileipellis and its smaller spores (Noordeloos 1987, 1988a, 1992, 2004, 2012; Noordeloos and Hausknecht 1993). Finally, E. minutum (P. Karst.) Noordel. is characterised by a strongly translucently striate pileus, often with a slight depression in the centre, subisodiametrical spores, and growth in moist grasslands, on damp places with Alnus, Fraxinus, Betula and Salix (Noordeloos 1992, 2004).

Entoloma calabrum (holotype) a Basidiomes b Pileus c Stipe. Scale bars: 1 cm
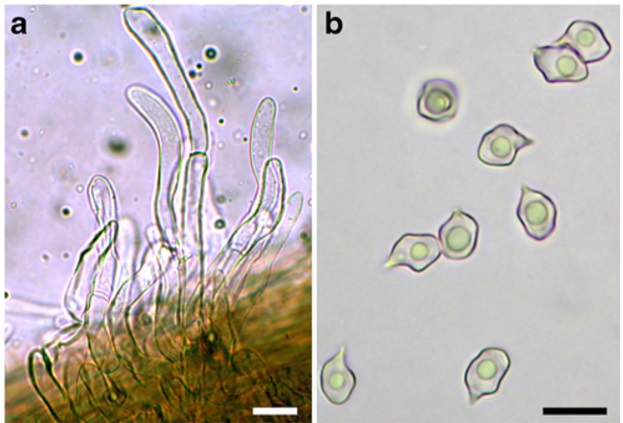
Entoloma calabrum (holotype) a Caulocystidia b Spores. Scale bars: 10 μm